

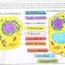
Każdy żywy organizm, pomijając kontrowersyjne wirusy, składa się, z co najmniej jednej komórki. Różnorodność życia na Ziemi jest ogromna, jego istota jednak zawsze pozostaje taka sama. Jest nią komórka. Począwszy od prostych jednokomórkowych organizmów takich jak pierwotniaki, a na człowieku skończywszy, to komórka zawsze stanowi podstawową cegiełkę budującą życie. Tworząc organizm komórka wyspecjalizowuje się do pełnionych przez siebie funkcji. To różnicowanie ujawnia się wyraźnie już u pierwszych tkankowców, ale nawet pojedyncza komórka tworząca organizm pierwotniaka, potrafi elastycznie dostosowywać się do zmian środowiska i specjalizować w kierunku tych funkcji życiowych, które przynoszą największe korzyści i dają organizmowi największe szanse na przeżycie. Taka specjalizacja prowadzi do tego, iż pomimo zachowania podstawowych cech budowy, poszczególne rodzaje komórek mogą zasadniczo różnić się od siebie.
Budowa komórki i funkcje jej składników
Wszystkie komórki wytworzyły specyficzną barierę chroniącą je przed negatywnym wpływem środowiska, tj. błonę komórkową, zbudowana z białek i lipidów. Jej budowę porównuje się do płynnej mozaiki. Wśród tłuszczowców dominują fosfolipidy, choć równie często występują glikolipidy, sfingolipidy i cerebrozydy (związki będące połączeniem glikolipidów ze sfingozyną wchodzącą w skład sfingolipidów). Tłuszczowce tworzą podwójną warstwę, tak skonstruowaną, że ich hydrofilowe końce znajdują się na zewnątrz, a hydrofobowe części skierowane są do wewnątrz. Pomiędzy lipidami znajdują się białka, które mają możliwość zmiany miejsca w błonie – przemieszczania się. Białka budujące błonę komórkową dzielimy na białka powierzchniowe oraz na białka integralne, które wbudowane w podwójny zrąb lipidowy mogą tworzyć kanały przenośnikowe. U zwierząt na powierzchni błony występują przyłączone do niektórych białek reszty węglowodanowe, tzw. glikokaliks. Chronią one błonę przed enzymami trawiennymi, np. w jelitach oraz pozwalają układowi odpornościowemu na odróżnienie komórek własnego organizmu od ciał obcych. Błona komórkowa występuje zarówno u zwierzą t jak i u rośli. Należałoby tu podkreślić, iż nie jest ona strukturą statyczną. Ulega ona ciągłym zmianom i przebudowom.
U roślin występuje dodatkowo ściana komórkowa. Ma ona charakter statyczny. Jej podstawowymi składnikami budulcowymi są: pektyny, celuloza, hemiceluloza, lignina. Ściana wtórna powstaje ze ściany pierwotnej na skutek:
- inkrustacji (wysycania wewnątrz ściany) ligniną (proces ten nazywamy drewnieniem), lub węglanem wapnia lub krzemionką
- adkrustacji (odkładania związków na powierzchni ściany pierwotnej) tłuszczowcami, np. suberyną (korkowacenie) lub polisacharydami
Ściana komórkowa zawiera tzw. składniki szkieletowe (40% s.m.) stanowiące podstawę jej konstrukcji (tj. celuloza – większość roślin, chityna – grzyby, ksyloza lub mannoza – niektóre glony) oraz tzw. składniki podłoża (białka, pektyny). Ściana komórkowa zawiera również znaczne ilości wody.
Wszystkie komórki (pomijam martwe komórki, np. komórki tworzące ksylem) zawierają cytoplazmę będącą swoistym środowiskiem wewnątrzkomórkowym. Cytoplazma jest roztworem białek (np. enzymatycznych, strukturalnych), lipidów i cukrów, jonów (np. Na+, Mg2+, K+, P+), kwasów tłuszczowych oraz innych związków, w którym woda stanowi fazę rozpuszczającą. Cytoplazma wykonuje aktywny ruch. U Eucaryota wyróżniamy trzy rodzaje ruchu cytoplazmy:
- ruch rotacyjny - wokół jednej centralnej wodniczki
- ruch cyrkulacyjny – wokół wielu mniejszych wodniczek
- ruch pulsacyjny – wokół wielu mniejszych wodniczek, w zmiennym kierunku
Cytoplazma jest układem wielofazowym, łatwo zmienia swój stan skupienia. Raz zachowuje się jak substancja półpłynna (sol), a raz jak półstała (żel). Proces pozwalający na przejście z solu w żel nazywamy koagulacją, odwrotny zaś peptyzacją.
Cytoplazma nie jest strukturą jednorodną. U Eucaryota podzielona jest siateczką wewnątrzplazmatyczną: szorstką (ERg) – połączoną z rybosomami; gładką (ERa) – gładką – nie połączoną z rybosomami. Siateczka wewnątrzplazmatyczna ma za zadanie umożliwić uzyskanie optymalnego środowiska dla wielu reakcji chemicznych. Każda reakcja chemiczna wchodząca w skład jednego ze szlaków metabolicznych posiada inne optymalne warunki. Siateczka oddziela miejsca, w których zachodzą jedne reakcje, od pozostałych. Ponadto ERg bierze udział w syntezie białek. ERa natomiast wspomaga syntezę lipidów i steroli. Siateczka wewnątrzplazmatyczna uczestniczy również w wielu szlakach metabolicznych, w tym w unieczynnianiu toksyn (w komórkach wątroby). W komórkach mięśniowych – sarkomerach występuje tzw. siateczka sarkoplazmatyczna. Uczestniczy ona w przekazywaniu impulsów nerwowych warunkujących skurcz mikrofibryli – filamentów aktynowych i miozynowych.
W cytoplazmie występują również białka o charakterze włókien, tzw. mikrofilamenty oraz mikrotubule (rurki). Mikrofilamenty umożliwiają aktywny ruch cytoplazmy, mikrotubule zaś mogą tworzyć wrzeciono kariokinetyczne, występujące przy podziale komórki.
Wszystkie wymienione do tej pory składniki stanowią zamknięte środowisko komórki, w którym umieszczone zostają poszczególne organella komórkowe, organella umożliwiające poprawną pracę komórki, stanowiącą o poprawnym funkcjonowaniu całego organizmu.
Najważniejszym organellum komórki jest jądro. To ono stanowi swoiste centrum dowodzenia i kontroli procesów życiowych. Materiał genetyczny zawarty w jądrze jest zbiorem informacji, programem, który jest realizowany przez poszczególne jednostki funkcjonalne wchodzące w skład komórki. Jądro wykształciło się u organizmów Eucariotycznych. Właściwie posiadanie jądra stanowi podstawowy element wyróżniający i pozwalający klasyfikowanie organizmów. Podstawą klasyfikacji jest podział na organizmy bezjądrowe – Procariota, będące organizmami jednokomórkowymi i znacznie prostszymi w budowie oraz organizmy posiadające jądro – Eucariota, charakteryzujące się znacznie bardziej złożoną budową. Organizmy eucariotyczne w toku ewolucji zdominowały zarówno świat roślin jak i zwierząt. Ich różnorodność jest zdumiewająca. Od prostych jednokomórkowych organizmów bakteryjnych, glonów, do tak skomplikowanych jak gromada ssaków. Wykształcenie jądra było krokiem koniecznym. Większa komplikacja organizmów wiązała się ze zwiększoną ilością DNA, które musiało zmieścić się w takiej samej, jeśli nie mniejszej objętości. Upakowanie w postaci luźnej nici chromatynowej okazało się niewystarczające. Poza tym, specjalizacja komórek w obrębie organizmu, tworzenie tkanek, wymagało wykorzystywania tylko nieznacznego fragmentu DNA, przez poszczególne komórki. Cała reszta, niepotrzebna, musiała zostać unieczynniona, zablokowana, dla enzymów transkrypcji DNAmRNA (polimeraza RNA). Konieczność ta odzwierciedlona jest w budowie i ułożeniu poszczególnych genów w DNA. U organizmów procariotycznych, geny kodujące białka – enzymy wchodzące w skład jednego szlaku metabolitycznego ułożone są liniowo. I tak transkrybująca polimeraza RNA przepisuje na mRNA całą informację potrzebną do późniejszego procesu translacji i syntezy odpowiednich białek. Fragment taki nosi nazwę operonu. Jest to pewnego rodzaju ułatwienie umożliwiające przyspieszenie procesów metabolicznych u organizmów procariotycznych. Poza tym u procariota cały DNA zorganizowany jest w jedną kulistą cząsteczkę, tzw. chromosom bakteryjny. Jest on dostępny dla procesów transkrypcyjnych prawie na całej długości. Jego ilość i budowa nie pozwala na różnicowanie się komórek, stąd wszystkie organizmy bezjądrowe występują w postaci jednokomórkowej. U jądrowych uproszczenie takie nie jest możliwe, a to ze względu na znacznie większą ilość informacji, a co za tym idzie większą komplikację w ułożeniu genów kodujących białka. Transkrypcji ulega tu, fragment odpowiedzialny za syntezę jednego, konkretnego białka. DNA posiada fragmenty kodujące – egzony, oraz fragmenty nie kodujące, pełniące najprawdopodobniej funkcję stabilizującą ogromnej, podwójnej helisy. Helisa ta zostaje nawinięta na specjalne białka, tzw. oktamery histonowe. Białek histonowych jest 5 rodzajów. Cztery z nich występują w parach tworząc oktamer, piąty występuje oddzielnie i pełni funkcje stabilizujące. Fragment taki nosi nazwę nukleosomu. Fragmenty te zwinięte są w spiralę i tworzą solenoid, który z kolei w całości skręca się tworząc domeny, które spinane są białkami niehistonowymi. Cała ta struktura tworzy osobne twory tzw. chromosomy. U zdrowego człowieka występuje ich aż 46. Tak upakowany materiał genetyczny wchodzi w skład jądra. Jąderko odgrodzone jest od reszty komórki barierą, tzw. błoną jądrową. Błona jądrowa od wewnątrz jest gładka od zewnątrz często przechodzi w ERg. W błonie jądrowej występują liczne pory, których wielkość i ilość zależy od poziomu metabolizmu komórki. Im jest on wyższy tym więcej i większe pory. Umożliwia to sprawniejsze wydostawanie się mRNA z jądra. Należy tutaj jeszcze wspomnieć, iż u Eucariota w przeciwieństwie do organizmów bezjądrowych proces transkrypcji jest oddzielony od procesu translacji. Pierwszy następuje wewnątrz jądra drugi zaś w obrębie cytoplazmy. Wnętrze jądra wypełnione jest tzw. kariolimfą. Jest to wodny roztwór białkowy zawierający enzymy m.in. polimerazę DNA i RNA.
W niektórych przypadkach na skutek specjalizacji komórek jądro ulega zanikowi. Występuje to m.in. w komórkach transportujących, np. erytrocyty, rurki sitowe. Umożliwia to znaczne ograniczenie metabolizmu komórki. W przypadku komórek sitowych brak jądra umożliwia łatwiejszy ruch cytoplazmy transportującej asymilaty. Brak jądra wpływa jednak niekorzystnie na żywotność komórek. I tak w przypadku erytrocytów wynosi około 100 dni a w przypadku rurek sitowych 1 rok.
Organellami umożliwiającymi wykonywanie poleceń „wydanych przez jądro” są rybosomy. To one przeprowając proces translacji tworzą gotowe produkty, których schematy zostały zapisane w DNA. Wyróżniamy dwa rodzaje rybosomów. Małe występują u procariota oraz w organellach autonomicznych, posiadających własne DNA (mitochondria i plastydy). Duże zaś są charakterystyczne tylko dla jądrowych. Niezależnie od rodzaju każdy rybosom składa się z dwoch jednostek: mniejszej i większej. W nieczynnym rybosomie podjednostki te występują oddzielnie. Gdy jednak następuje konieczność uruchomienia aparatu translacyjnego podjednostki te łączą się tworząc jedną całość. Połączenie to warunkuje odpowiednie stężenie jonów Mg+. Rybosom składa się z dwóch typów białek: kwaśnych – pełniących funkcję enzymatyczną i zasadowych – strukturalnych. Sam rybosom nie warunkuje jednak odpowiedniego przebiegu procesu translacji. „Współpracuje” on z cząsteczkami tRNA, które odpowiedzialne są za transport odpowiednich aminokwasów w obręb rybosomu. Cząsteczki tRNA posiadają tzw. pętlę antykodonową, dzięki której możliwe staje się odróżnienie odpowiednich nukleotydów i dopasowanie odpowiednich aminokwasów. W rybosomie, zgodnie z zasadą komplementarności, tRNA łączy się z łańcuchem mRNA. W podjednostce dużej jest miejsce na dwie cząsteczki tRNA. Gdy obydwa „doki” zostaną zapełnione enzym wchodzący w skład dużej jednostki, katalizuje wytworzenie wiązania peptydowego, pomiędzy aminokwasami. Wkrótce potem pierwsza cząsteczka tRNA odłącza się od „swojego aminokwasu i uwalnia się do cytoplazmy. Drugi tRNA wraz z dołączonym, w tym wypadku, już dipeptydem, przesuwa się w miejsce pierwszego, a jego miejsce zajmuje kolejna cząsteczka tRNA z kolejnym aminokwasem. Wytwarza się kolejne wiązanie peptydowe pomiędzy sąsiednimi aminokwasami. Proces ten przebiega aż do zrealizowania całego programu mRNA, czyli aż do wystąpienia trójki nonsensownej (UUA, UGA, UAG) i nosi nazwę elongacji translacyjnej.
Podziały komórkowe
Wszystkie, komórki posiadają pierwotną umiejętność dzielenia się. Na drodze mitozy, z każdej komórki mogą powstać dwie, identyczne pod względem zawartego w nich DNA. Tak podział pozwala na rozwój i rozrost organizmu. Ponadto u organizmów jednokomórkowych pozwala na szybkie i wydajne rozmnażanie. Podstawowymi założeniami procesu mitotycznego jest niezmienność ilości i struktury DNA komórek potomnych w porównaniu do komórki macierzystej. Mitoza składa się z jednego cyklu podziałowego. Zachodzi zwykle w komórkach somatycznych organizmów. 70% do 90% czasu całego cyklu mitotycznego stanowi interfaza, która dzieli się na trzy okresy. Okres G1 będący następstwem poprzedniej telofazy. W nim to komórka koncetruje się na odbudowie i powrocie do stanu z przed podziału. Dominują tu procesy anabolityczne. Następuje wzrost produkcji fosfolipidów, związany z koniecznością odbudowy błony komórkowej. Również wysoki jest poziom syntetyzowanych białek spowodowany wzrostem aktywności transkrypcyjnej i translacyjnej. Okres G1 może przejść bezpośrednio w okres S lub od razu w okres G2. W fazie S następuje intensywna replikacja DNA i związana z nią dekondensacja chromatyny. Natomiast w okresie G1 dochodzi do podziału organelli samoreplikujących się (m.in. mitochondria, chloroplasty) oraz z powrotem narasta intensywność procesów transkrypcyjnych i translacyjnych związanych z tworzeniem białek wrzeciona kariokinetycznego (tubulina). Okres G1 stanowi koniec interfazy. Po niej następuje właściwa kariokineza, kto rej towarzyszy podział cytoplazmy. Rozpoczyna się profazą, w której to chromatyna ulega kondensacji w chromosomy (każdy składający się z dwóch chromatyd). Chromosomy gromadzą się w środkowej części komórki, a błona jądrowa ulega pęknięciu. Następuje okres metafazy, w której to do każdego chromosomu w miejscu tzw. przewężenia pierwotnego przyłącza się mikrotubula wrzeciona podziałowego. Następuje uporządkowanie chromosomów. Tworzą one teraz tzw. płytkę metafazalną. W momencie pęknięcia ostatniego cetromeru rozpoczyna się anafaza. Chromosomy potomne odciągane są przez wrzeciona popychając przed sobą organella komórkowe. Niedługo później rozpoczyna się telofaza. Rozpoczyna się procesem cytokinezy. Odtworzeniu ulega otoczka jądrowa a chromosomy ulegają despiralizacji. Przedstawiony w skrócie, powyżej proces mitozy warunkuje wzrost organizmu. Różni się on diametralnie od procesu mejotycznego.
Mejoza stanowi proces warunkujący różnicowanie się organizmów. W odróżnieniu od mitozy składa się z dwóch cykli podziałowych, pomiędzy którymi nie następuje replikacja DNA. W jej wyniku powstają cztery komórki potomne o zredukowanej o połowę ilości informacji genetycznej. Zachodzi w komórkach rozrodczych. Ze względu na moment, w którym zachodzi wyróżniamy trzy rodzaje mejozy: pregamiczna – następuje przed połączeniem się gamet, przed zapłodnieniem; zachodzi u organizmów diploidalnych; postgamiczna – następuje po zapłudnieniu – u organizmów haploidalnych; trzeci rodzaje mejozy jest mejoza pośrednia, która zachodzi w diploidalnym sporoficie podczas powstawania zarodników, z których to wyrasta haploidany gametofit.
Interfaza mejozy jest bardzo podobna do mitotycznej. Składa się jednak z dwóch okresów G1 i S, z tymże okres G1 jest identyczny jak ten występujący w mitozie. Mejotyczny okres S jest trzy razy dłuższy od mitotycznego. Następuje w nim replikacja DNA oraz syntetyzowana jest tubulina. Poprzedza on bezpośrednio kariokinezę.
W mejozie – wyróżniamy dwa cykle podziałowe, każdy złożony z czterech faz.
I – cykl podziałowy – heterotypowy – redukcja chromosomów z 2n do n i ilości DNA z 4c do 2c; następuje tu proces crossing-over.
a) profaza I
- leptoten – pojawienie się cienkich, gładkich i splątanych nici chromosomów (kondensacja domen i obudowanie ich białkami szkieletowymi
- zygoten – koniugacja chromosomów homologicznych (boczne pętle niecałkowicie zreplikowanych odcinków z-DNA); ułożona równolegle para tworzy biwalent; synteza enzymu endonukleazy
- pachyten – kondensacja chromosomów w biwalentach; zachodzi crossing-over – powstają chiazmy
- diploten – oddzielenie chromosomów homologicznych; jedynymi ptk. Kontaktu zostają chiazmy
- diakineza – przesuwanie chiazm wzdłuż biwalentów ku ich końcom; skręcanie i grubienie chromosomów; stadium kontrakcji
b) metafaza I – włókna wrzeciona kariokinetycznego doprowadzają do rozerwania biwalentów – pęknięcie chiazm
c) anafaza I – odciąganie chromosomów homologicznych do przeciwległych biegunów komórki
d) telofaza I – odtwarzanie otoczki jądrowej
II – cykl podziałowy – homotypowy
a) profaza II – tworzenie włókien wrzeciona kariokinetycznego, zanikanie otoczki jądrowej i jąderek
b) metafaza II – rozrywanie centromerów przez włókna wrzeciona kariokinetycznego
c) anafaza – do przeciwległych biegunów wędrują chromosomy potomne – rozpoczęcie cytokinezy
d) telofaza II – odtwarzanie otoczki jądrowej, jąderek, despiralizacja chromosomów
Proces mejozy jest bardzo istotny z ewolucyjnego punktu widzenia, szczególnie ze względu na niezależną segregację chromosomów oraz proces c.o. Umożliwia on w drodze w drodze doboru naturalnego, przystosowanie się organizmów do nowych warunków środowiskowych, a co za tym idzie ich ciągły rozwój.
Występuje jeszcze jeden rodzaj podziału komórkowego. Nie ma on jednak tak dużego znaczenia jak proces mitozy czy mejozy. Jest to proces polegający na bezpośrednim podziale jądra komórkowego – tzw. amitoza. Nie jest to proces dokładny, jeżeli chodzi o zachowanie zgodności informacji genetycznej komórek potomnych z komórką rodzicielską. Występuje w jądrach posiadających wiele kopii DNA – poliploidalnych. Charakterystyczny dla orzęsków (makronukleus). Występuje również w komórkach zmutowanych i starych.
Jednymi z najważniejszych organellów komórkowych, które jednocześnie są charakterystyczne i pozwalają odróżnić rośliny od zwierząt są odpowiednio u roślin chloroplasty a o zwierząt mitochondria.
Chloroplasty należą do szerszej grupy plastydów, pełniących o roślin różnorakie funkcje. Wszystkie posiadają własny DNA oraz aparat translacyjny. Większość posiada również rozbudowany system błon wewnętrznych zanurzonych w stromie. Wszystkie rodzaje plastydów powstają z tzw. proplastydów i różnicują się pod wpływem oddziaływań środowiskowych, oraz ze względu na tkankę, do której należą komórki, w których skład wchodzą. I tak wszelkie barwne plastydy – chloroplasty, chromoplasty mogą przekształcić się w bezbarwne etioplasty, gdy pozbawi się do nich dopływu światła. Plastydy mogą sprawować funkcję organelli zapasowych. Leukoplasty, mające słabo rozwinięty system błon wewnętrznych gromadzą w swoim wnętrzu skrobię. Gdy całkowicie zostaną wypełnione przez skrobie przechodzą w amyloplast. Występują w korzeniach, mogą jednak pod wpływem światła przekształcić się w chloroplasty. Mogą również zamiast skrobi gromadzić białka zapasowe, nazywane wtedy są proteinoplastami. Chloroplasty odpowiedzialne są za tworzenie, w procesie fotosyntezy, energii potrzebnej do funkcjonowania komórki. Energia gromadzona jest w ATP na skutek fosforylacji fotosyntetycznej, która odbywa się w błonach chloropalstów, tzw. kwantosomach.
U zwierząt, wraz z przejściem z samożywności na cudzożywny sposób odżywiania, pojawiły się mitochondria. Należą one również do organelli autonomicznych, ponieważ posiadają własny DNA tzw. mitochondrialny oraz rybosomy. Podobnie jak chloroplasty otoczone są podwójną błoną. Pomiędzy błonami występuje przestrzeń perymitochondrialna. Wewnętrzna błona wpukla się do wewnątrz tworząc grzebienie mitochondrialne, na których to zachodzi proces fosforylacji oksydacyjnej. Wnętrze tych organelli, porównywalnych często do elektrowni komórkowych, wypełnione jest tzw. matrix mitochondrialnym. W roztworze tym rozpuszczone są enzymy katalizujące przebieg m.in. szlaku b-oksydacji i cyklu Krebsa.
Specyficzną funkcję pełni w komórce Aparat Golgiego. Występuje on tylko u organizmów eukariotycznych. Składa się z diktiosomów, będących mniej lub bardziej spłaszczonymi woreczkami zbudowanymi z gładkich błon biologicznych, ułożonych na sobie. Aparat Golgiego jest silnie związany retikulum endoplazmatycznym. Uczestniczy w rozprowadzaniu związków w obrębie komórki, syntezie błony komórkowej. Dokonuje również syntezy polisacharydów, mukosacharydów oraz umożliwia połączenie węglowodanów z proteinami. Uczestniczy również w wydzielaniu wielu substancji poza komórkę w postaci egzocytozy.
Można by powiedzieć, że organellami pełniącymi w komórce funkcję odwrotną niż aparaty Golgiego są lizosomy i mikrociałka. Są one syntetyzowane przez aparaty Golgiego. Pełnią one funkcję trawienną. Posiadają w swoim wnętrzu dezaktywowane enzymy hydrolityczne. Dzięki nim mogą utleniać substraty, uczestniczą w mobilizacji rezerw tłuszczowych. Współdziałają również w procesach fotooddychania. W połączeniu z fagosomami tworzą lizosom wtórny (uczestniczą w procesach endocytoz). Mikrociałka, są tworami posiadającymi szereg enzymów z grupy oksydaz L- i D-aminokwasowych (peroksysomy) oraz enzymy –oksydacji (glioksysomy). Peroksysomy pełnią ważną funkcję w procesie rozkładu toksycznego H2O2. Występowanie w peroksysomach enzymów oksydaz L-aminokwasowych stanowi swego rodzaju skamieniałość ewolucyjną. Enzymy te nie odgrywają już dzisiaj żadnej roli w procesach życiowych, gdyż wszystkie żywe organizmy posiadają tylko D-aminokwasy. Najprawdopodobniej kiedyś D-aminokwasy występowały obok L- stąd prawdopodobnie pochodzi ta pozostałość w postaci enzymów.
U Eucariota występuje jeszcze jedna struktura, mianowicie wakuola. Występuje zarówno u roślin jak i u zwierząt, z tą tylko różnicą, że u tych pierwszych występuje zazwyczaj jedna (lub kilka) dużych wakuoli, a u zwierząt jest ich znacznie więcej, mniejszych. Tonoplast otoczony jest błoną wakuolarną. Jego wnętrze wypełnione jest sokiem wakuolarnym, składającym się w głównej mierze z wody (90%). Rozpuszczone są w niej różnego rodzaju jony. Mogą również występować związki nieorganiczne, które wytrąciły się z roztwory (np. kryształy szczawianu wapnia). Gromadzą się tutaj również metabolity (glikozydy, alkaloidy, garbniki) oraz aminokwasy, białka, kwasy organiczne. Wakuola na skutek całkowitego wypełnienia przez białko może ulec przekształceniu w tzw. ziarno aleuronowe. Po rodzaju składników gromadzonych w wakuoli możemy rozróżnić pełnione przez nią funkcje. I tak woda w niej zgromadzona pozwala na utrzymanie właściwego uwodnienia komórki. Metabolity – magazynowanie zbędnych produktów przemiany materii. Ponadto u pierwotniaków wakuola może przekształcić się w tzw. wodniczkę tętniącą, odpowiedzialną za usuwanie nadmiaru wody gromadzącej się w organizmie na skutek osmozy. W połączeniu z lizosomami wakuola, tworzy lizosom wtórny, czyli tzw. wodniczkę pokarmową. Pełni to ważną rolę w procesach odżywiania u pierwotniaków.
Wszystkie, przedstawione powyżej składniki komórki, stanowią jej nierozłączną całość. Niektóre z nich występują tylko u roślin inne zaś są charakterystyczne wyłącznie dla organizmów zwierzęcych. Jedne powstały w toku ewolucji dopiero u Eucariota inne występowały już u prymitywnych bakterii. Na temat ewolucji komórek jest wiele teorii. Jedna z nich mówi, że zarówno mitochondria jak i chloroplasty były kiedyś oddzielnymi organizmami. Jedne wyspecjalizowały się w syntezie związków potrzebnych im do życia przy pomocy promieniowania świetlnego drugie natomiast otrzymywały ja przeprowadzając procesy utleniania związków i rozkładu związków bardziej złożonych do prostszych. Wszystkie te odmienne strategie, począwszy od foto-, przez chemosyntezy aż do całkowitej cudzożywności, dały początek całej grupie nowych organizmów. Uważa się, że zarówno mitochondria jak i chloroplasty weszły w ścisłą symbiozę mutualistyczną z pierwotnymi organizmami jednokomórkowymi. Jedne rozwinęły się w dzisiejsze rośliny drugie zaś dały początek zwierzętom. Ogniwem je łączącym, ma być według współczesnych biologów euglena zielona (Euglena viridis) będąca prymitywnym organizmem jednokomórkowym łączącym w sobie cechy charakterystyczne zarówno dla roślin, jaki i dla zwierząt.
W drodze ewolucji poszczególne rodzaje komórek ulegały coraz to większej specjalizacji. Organizmy jednokomórkowe rozpoczęły życie w koloniach (skrętnice – Spirogyra). Funkcje poszczególnych komórek ulegały różnicowaniu (toczek – Volvox). Pojawiły się pierwsze nibytkanki – organizmy plektenchymatyczne (morszczyn), poźniej tkanki (mszaki, paprotniki). W królestwie zwierząt z organizmów jednokomórkowych powstały pierwsze wielokomórkowe. W drodze ewolucji po bezkręgowcach nastąpiły strunowce. Coraz wyżej uorganizowane. W końcowej fazie pojawiły się ssaki a na końcu człowiek. Różnorodność organizmów jest olbrzymia. Wszystkie składają cię jednak z podstawowych cegiełek życia – komórek. Możemy sobie zadać pytanie czy tak naprawdę pojedyncza komórka wątroby różni się zasadniczo od neuronu, a ten czy odbiega zasadniczo od budowy komórki pierwotniaka. Przecież tak samo jak wszystkie inne posiada informację zawartą w DNA. Różni się tylko sposobem jej upakowania i ilością. Gdy oglądamy pod mikroskopem neuron i komórkę wątroby różnią się one diametralnie budową. Jest to jednak głównie różnice w budowie zewnętrznej, spowodowana przystosowaniem do pełnionych prze siebie funkcji. Komórka wątroby nie musi się kontaktować i wymieniać impulsów elektrycznych z pozostałymi. Nie musi również wytwarzać skomplikowanych przekaźników neurochemicznych, aby pobudzić działanie „sąsiadki”. Musi natomiast radzić sobie z unieczynnianiem ogromnej ilości toksyn, zatruwających nasz organizm m.in. po to, aby każdy neuron w organizmie mógł poprawnie funkcjonować. Funkcjonowanie organizmu polega na ciągłe współpracy wszystkich komórek wchodzących w jego skład. Im bardziej skomplikowany organizm tym ta współpraca musi być dokładniejsza i lepiej kontrolowana. Wiadomo przecież, że tak samo jak nie ma maszyny, która spełniałaby wszystkie możliwe czynności, tak nie możliwe jest stworzenie komórki, która mogłaby wypełniać wszystkie skomplikowane czynności, jakich natura wymaga od organizmu, tylko po to, aby mógł przetrwać.
Informacja od autora
Praca ta jest próbną pracą maturalną. Została oceniona na 4. W uzasadnieniu oceny stwierdzono , że „w stosunku do bardzo rozbudowanej treści dotyczącej jądra pojawiło się zbyt mało informacji dotyczącej odżywiania komórek auto- i heterotroficznych oraz oddychania tlenowego i beztlenowego. Brak informacji dotyczącej wrażliwości komórki (teoria Schleiden’a i Schwann’a).